Le terme ‹ morphologie › se rapporte à deux branches de la Biologie, une qui traite de la forme et de la structure des organes ou d'autres parties des organismes, et l'autre de la forme et la structure d'un organisme dans son entier.
De la même façon, la table MORPHOLOGY vise à accomplir deux objectifs connexes :
i. fournir des descriptions concises standardisées des poissons inclus dans FishBase pour les rendre comparables ; et
ii. permettre l'identification rapide des espèces grâce aux caractères indiqués dans (i).
Chez les poissons, les principaux caractères utilisés pour la description et l'identification sont des attributs descriptifs, qui font référence aux caractères distinctifs (par exemple, la forme de la nageoire caudale), des mesures morphométriques, qui font référence à des variables numériques continues (par exemple, la longueur de la tête par rapport à la longueur du corps) ou des comptages méristiques, qui font référence à des variables numériques discontinues (par exemple, le nombre des rayons mous et épineux pour la nageoire dorsale).
La table MORPHOLOGY contient les caractères textuels dans des champs à choix multiples, et les caractères morphométriques et méristiques dans des champs numériques. Ce sont principalement les caractères méristiques qui sont utilisés pour une identification rapide, suivant le processus établi pour les bases de données par Froese et Papasissi (1990). La structure de la table MORPHOLOGY et les choix disponibles dans les champs résultent d'une étude approfondie d'ouvrages majeurs en ichtyologie (par exemple, Lagler et al. 1977) et de la consultation de nombreux collègues. Quelques-uns des termes employés dans la table sont très spécialisés, leur définition se trouve dans le Glossaire.
Les champs
La table MORPHOLOGY comprend 67 champs à choix multiple, 79 champs numériques et plusieurs champs de remarques. D'une part les champs à choix multiple présentent à l'utilisateur des caractères prédéfinis pour une partie ou une caractéristique du corps (par exemple, Cross section [Section transversale] : circular [circulaire] ; oval [ovale] ; compressed [comprimée] ; flattened [aplatie] ; angular [anguleuse] ; others [autres] (voir Remarks [Remarques])). Le nombre de ces choix a été maintenu à un minimum, en ne retenant que ceux qui se rapportent aux formes les plus communes.
Dans la plupart des cas, un choix ‹ other › (see Remarks) [autres (voir Remarques)] permet d'indiquer chez certaines espèces, des caractères ou des conformations aberrantes d'une partie du corps qui sont alors détaillés dans Remarks [Remarques].
D'autre part, la majorité des caractères morphométriques et méristiques, sont présentés sous forme d'intervalle, deux champs numériques contenant l'un la limite inférieure, l'autre la limite supérieure. Quand un intervalle ou plusieurs valeurs ont été données dans la littérature, mais qu'un seul champ est disponible, comme les proportions du corps, la moyenne des valeurs disponibles est indiquée.
Le champ Remarks [Remarques] contient la description des caractères qui ne sont pas inclus dans la structure ou les choix des champs, ou qui requièrent des descriptions plus détaillées. Dans ces champs, les caractéristiques distinctives sont mises en exergue, de même si ces caractéristiques peuvent être rencontrées chez des espèces apparentées. Les variations de couleur (ontogénétiques, sexuelles et géographiques) sont aussi commentées quand l'information est disponible.
Au fur et à mesure que le nombre d'espèces s'est accru dans FishBase, remplir plus de 140 champs de la table MORPHOLOGY pour toutes les espèces est devenu trop prenant. Nous avons décidé de réduire le nombre de champs ‹ actifs › et de ne renseigner régulièrement que ceux qui correspondent aux caractères généralement décrits dans les travaux taxinomiques (méristiques standard et diagnostiques). Ainsi, une description minimale a été saisie pour tous les poissons osseux du Japon et de la Colombie Britannique, et tous les poissons marins de Micronésie et d'Afrique australe (Smith et Heemstra 1986). De même, toutes les familles traitées dans les catalogues FAO ou dans la série Indo-Pacific Fishes de Randall sont complètes. Nous prévoyons de compléter et de vérifier la table MORPHOLOGY par famille (voir Encadré 1, ce volume) et en utilisant les travaux faunistiques comme Skelton (1993) Freshwater Fishes of Southern Africa.
Utilisation
Une utilisation importante des informations contenues dans la table MORPHOLOGY est l'identification rapide des poissons (voir ‹ Identification rapide ›, ce volume). La procédure préprogrammée actuelle exige un minimum d'informations comme critères de recherche, viz. :
La procédure recherche les espèces qui correspondent aux critères fournis par l'utilisateur et en affiche la liste, typiquement moins de 10 espèces pour une même famille. L'utilisateur peut ensuite afficher des illustrations et les descriptions morphologiques complètes pour achever ou vérifier une identification. Cette procédure de recherche fonctionne aussi si un ou plusieurs des critères manquent. Dans ce cas, la liste d'espèces est plus longue. Noter cependant que la table MORPHOLOGY ne contient des données que pour 6 300 espèces environ et n'est complète que pour quelques zones ou familles (voir ci-dessus). Le degré de complétude des informations disponibles varie de très insuffisant, comme pour Pellona castelnaeana, à presque complet, comme pour Lutjanus biguttatus. De plus, la vérification des données n'est pas exhaustive et certaines peuvent donc être erronées.
Les rapports
Les procédures préprogrammées pour imprimer des synopsis et des résumés sur les espèces utilisent les informations de la table MORPHOLOGY. Par exemple, la procédure pour imprimer un synopsis d'espèce extrait les données de Additional Characters [Caractères supplémentaires] qui contient des descriptions diagnostiques, le nombre de rayons aux nageoires dorsales et anales, et d'autres informations des tables SPECIES et STOCKS. Elle imprime un rapport complet des informations disponibles pour cette espèce, ainsi que toutes les références utilisées (voir ‹ Les rapports › ce volume).
Sources
Les données de la table MORPHOLOGY proviennent de tous les volumes des FAO Species Catalogues publiés jusqu'à aujourd'hui, de révisions taxinomiques, de livres faunistiques et d'articles scientifiques, entre autres Burgess (1978), Trewavas (1983), Allen (1985), Cohen et al. (1990), Lévêque (1990), Randall et al. (1990), Allen (1991), Myers (1991), et Heemstra et Randall (1993).
Comment y arriver
Cliquer sur le bouton Biology dans la vue SPECIES, puis sur le bouton Morphology & Physiology dans la vue BIOLOGY, et sur le bouton Morphology dans la fenêtre MORPHOLOGY & PHYSIOLOGY. Pour la procédure Quick Identification, cliquer sur le bouton Species dans la fenêtre MAIN MENU, et sur le bouton Quick Identification dans la fenêtre SEARCH BY …
Références
Allen, G.R. 1985. FAO species catalogue. Vol. 6. Snappers of the world. An annotated and illustrated catalogue of lutjanid species known to date. FAO Fish. Synop. 6(125) : 208 p.
Allen, G.R. 1991. Damselfishes of the world. Mergus Publishers, Melle, Germany. 271 p.
Burgess, W.E. 1978. Butterflyfishes of the world. A monograph of the Family Chaetodontidae. T.F.H. Publications, Neptune City. 832 p.
Cohen, D.M., T. Inada, T. Iwamoto et N. Scialabba. 1990. FAO species catalogue. Gadiform fishes of the world (Order Gadiformes). An annotated and illustrated catalogue of cods, hakes, grenadiers and other gadiform fishes known to date. FAO Fish. Synop. 10(125) : 442 p.
Froese, R., et C. Papasissi. 1990. The use of modern relational databases for identification of fish larvae. J. Appl. Ichthyol. 6 : 37-45.
Heemstra, P.C. et J.E. Randall. 1993. FAO species catalogue. Vol. 16. Groupers of the world. (Family Serranidae, Subfamily Epinephelinae). An annotated and illustrated catalogue of the grouper, rockcod, hind, coral grouper and lyretail species known to date. FAO Fisheries Synop. 16(125) : 382 p.
Lagler, K.F., J.E. Bardach, R.R. Miller, et D.R. May-Passino. 1977. Ichthyology. Deuxième édition. John Wiley and Sons, New York. 506 p.
Lévêque, C. 1990. Cyprinidae, p. 269-361. In C. Lévêque, D. Paugy et G.G. Teugels (éds). Faune des poissons d'eaux douces et saumâtres d'Afrique de l'Ouest. Tome I. Coll. Faune Tropicale n° XXVIII. Musée Royal de l'Afrique Centrale, Tervuren, et O.R.S.T.O.M., Paris. 384 p.
Myers, R.F. 1991. Micronesian reef fishes. Deuxième édition. Coral Graphics, Barrigada, Guam. 298 p.
Randall, J.E., G.R. Allen et R.C. Steene. 1990. Fishes of the Great Barrier Reef and Coral Sea. University of Hawaii Press, Honolulu, Hawaii. 506 p.
Skelton, P.H. 1993. A complete guide to the freshwater fishes of Southern Africa. Southern Book Publisher, South Africa. 388 p.
Smith, M.M. et P.C. Heemstra, Éditeurs. 1986. Smith's sea fishes. Springer Verlag, Berlin. 1047 p.
Trewavas, E. 1983. Tilapiine fishes of the Genera Sarotherodon, Oreochromis and Danakilia. British Museum of Natural History, London. 583 p.
Rainer Froese et Rodolfo B. Reyes, Jr.
La table VISION, consacrée à la vision des poissons et principalement aux pigments de la rétine de leurs yeux, contient 409 enregistrements pour 371 espèces extraits des travaux de Denton et Warren (1956), Munz (1964), Munz et McFarland (1973), Ali et Wagner (1975), et Hobson et al. (1981).
Les champs
Les auteurs précités ont montré que la Sensitivity
[Sensibilité] d'un œil de poisson est maximale à une
certaine longueur d'onde (![]() max).
Cette valeur en nm, et ses 95 % d'intervalle de confiance
(si disponible) constituent les données essentielles de cette
table.
max).
Cette valeur en nm, et ses 95 % d'intervalle de confiance
(si disponible) constituent les données essentielles de cette
table.
Un champ oui/non permet d'indiquer la présence d'autres pigments (comme dans le Tableau 3 de Hobson et al. 1981).
Un champ de remarques complète cette petite table.
Les utilisateurs de cette table devraient étudier les
articles précités pour les détails sur les méthodes
d'estimation de ![]() max.
max.
Statut
Mettre à jour la table VISION impliquera :
Les informations dans cette table peuvent être utilisées pour tester des hypothèses qui relient la physiologie et l'écologie des poissons, comme l'ont initié les travaux ci-dessous.
Comment y arriver
Cliquer sur le bouton Biology dans la vue SPECIES, puis sur le bouton Morphology & Physiology dans la vue BIOLOGY, et sur le bouton Vision dans la fenêtre MORPHOLOGY & PHYSIOLOGY.
Références
Ali, M.A. et H.J. Wagner. 1975. Visual pigments : phylogeny and ecology, p. 481-516. In M.A. Ali (éd.) Vision in fishes. New approaches to research. Plenum Press, New York & London.
Denton, E.J. et F.J. Warren. 1956. Visual pigments of deep-sea fish. Nature 4541 : 1059.
Hobson, E.S., W.N. McFarland et J.R. Chess. 1981. Crepuscular and nocturnal activities of Californian nearshore fishes, with consideration of their scotopic visual pigments and the photic environment. U.S. Fish. Bull. 79 : 1-30.
Munz, F.W. 1964. The visual pigments of epipelagic and rocky-shore fishes. Vision Res. 4 : 441-454.
Munz, F.W. et W.N. McFarland. 1973. The significance of spectral position in the rhodopsins of tropical marine fishes. Vision Res. 13 : 1829-1874.
Daniel Pauly
La plupart des poissons ont des petits cerveaux, du moins comparés aux vertébrés homéothermes (oiseaux et mammifères). Cependant, retenir cette caractéristique contre eux serait aussi absurde que d'inférer la valeur de différents groupes de personnes en mesurant la taille (mal évaluée) de leur cerveau (Gould 1981).
Plutôt, nous devrions nous rendre compte que la taille de leur cerveau a évolué sous la contrainte de leurs besoins, et nous devrions comparer la taille du cerveau entre espèces de poissons pour inférer ces besoins, donc leur niche (voir, par exemple, Bauchot et al. 1989). La base de données sur la taille du cerveau des poissons assemblée par Roland Bauchot et ses collaborateurs et mise gracieusement à disposition de FishBase permet des inférences de cette sorte. La suite décrit comment cette base de données a été créée d'après Bauchot et Bauchot (1986).
Plus de 2 800 cerveaux ont été disséqués pour plus de 900 espèces de poissons téléostéens (voir Fig. 43). De nombreux spécimens ont été capturés dans les zones tropicales et subtropicales : Îles Hawaï, Îles Marshall, Nouvelle-Calédonie, Australie (Queensland), Philippines, sud-ouest de l'Inde, Île Maurice, Île de la Réunion, Golfe d'Oman, nord de la Mer Rouge, Sénégal et Caraïbes ; mais aussi en France et dans l'Atlantique Nord. Tous les poissons ont été pesés avant l'enlèvement du cerveau et leur longueur totale et/ou standard mesurée. Les cerveaux ont été séparés de la moelle épinière au niveau des premiers nerfs vertébraux, les méninges et les vaisseaux sanguins enlevés, puis ils ont été tamponnés sur du papier buvard et pesés, et enfin conservés dans une solution de Bouin.
| Encadré 28. La taille du cerveau et la
consommation d'oxygène. Disposant d'un grand
ensemble de données sur la taille relative du cerveau,
nous avons été tentés de tester des hypothèses
évidentes. La figure 44 illustre une première
tentative de relier la table BRAINS avec d'autres
données physiologiques, ici la table OXYGEN. Les deux
ensembles de données contiennent la taille des
spécimens, fortement corrélée au poids dans les deux
cas. |

Fig. 43. Poids relatif du cerveau en fonction du poids corporel. Points clairs : enregistrements divers dans FishBase ; Points sombres : requins et raies, qui ont relativement de grands cerveaux, peut-être pour assurer leur activité d'électro-détection. Par contraste, 6 des points sous le nuage correspondent à des lamproies.
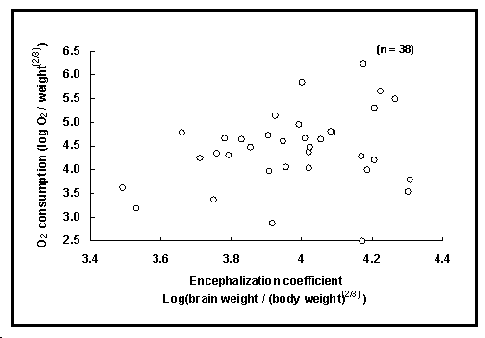
Fig. 44. Consommation d'oxygène en fonction du poids relatif du cerveau de 30 espèces de poissons. Voir Encadré 28 pour une interprétation de ce graphique.
Les juvéniles ayant un cerveau plus grand que les adultes relativement au poids corporel (Bauchot et al. 1979), les études comparatives n'ont principalement concerné que les adultes. Néanmoins, quelques mesures obtenues sur des séries allant des juvéniles à des grands adultes, permettent l'étude des changements ontogéniques de la taille du cerveau.
Les enregistrements pour un seul poisson ainsi obtenus sont présentés ici sous les noms courants d'espèce, et consistent en les éléments suivants :
Ces enregistrements sont présentés sous la forme d'un tableau par espèce, de une à 73 lignes.
Le travail suivant pour cette table sera l'intégration de plus de 200 enregistrements dont nous n'avons pas réussi à déterminer le nom valide dans FishBase. Un de nous (James Albert) du Department of Anatomy, Nippon Medical School, Tokyo, développe toujours cette table et a déjà ajouté 77 espèces représentant 18 nouvelles familles. Un article analyse cet ensemble augmenté de données (Albert et al., 1999). De plus, Mademoiselle Xiomara Chin de l'Institute of Marine Affairs, Trinidad & Tobago, a fourni les poids de cerveaux obtenus pendant son travail de thèse (Chin 1996).
Comment y arriver
Cliquer sur le bouton Biology dans la vue SPECIES, puis sur le bouton Morphology & Physiology dans la vue BIOLOGY, et sur le bouton Brains dans la fenêtre MORPHOLOGY & PHYSIOLOGY.
Remerciements
Nous remercions R. Bauchot et ses collaborateurs pour avoir confié à FishBase leurs précieux enregistrements, et J.-C. Hureau pour les transferts de format informatique. Nous remercions aussi Mlle X. Chin pour les 14 enregistrements des poids de cerveaux de poissons des Antilles.
Références
Albert, J.S., R. Froese, R. Bauchot et H. Ito. 1999. Diversity of brain size in fishes : preliminary analysis of a database including 1,174 species in 45 orders. In B. Séret et J.-Y. Sire (éds.) Proceedings of the 5th Indo-Pacific Fish Conference, Noumea, New Caledonia. pp. 647-656. Paris : Société Française d'Ichtyologie et IRD.
Bauchot, M.L. et R. Bauchot. 1986. Encephalization in tropical teleosts fishes and its correlation with their locomotory habits, p. 678-690. In T. Uyeno, R. Arai, T. Taniuchi et K. Matsuura (éds) Indo-Pacific Fish Biology : Proceedings of the Second International Conference on Indo-Pacific Fishes. Ichthyological Society of Japan, Tokyo.
Bauchot, R., M. Diagne et J.M. Ribet. 1979. Post-hatching growth and allometry of the Teleost brain. J. Hirnforsch. 20 : 29-34.
Bauchot, R., J.M. Ridet et M.-L. Bauchot. 1989. The brain organization of butterflyfishes. Environ. Biol. Fish. 25(1/3) : 205-219.
Chin, X. 1996. A photographic atlas of brains of common Caribbean reef fishes. University of South Florida, B.A. thesis. 62 p.
Gould, S.J. 1981. The mismeasure of man. W.W. Norton, New York. 352 p.
Daniel Pauly, Rainer Froese et James S. Albert
Le métabolisme est un processus physiologique qui reflète la dépense d'énergie des organismes vivants hétérotrophes, et de là, leurs exigences alimentaires. Le taux métabolique des poissons est généralement mesuré par leur taux de respiration, c'est-à-dire leur taux de consommation d'oxygène (voir Fig. 45). Les informations sur la consommation d'oxygène ne sont pas seulement utiles pour la physiologie comparative, mais aussi pour la pisciculture et la gestion des pêches. Elles fournissent des indices pour résoudre des problèmes associés à l'élevage ou au transport des poissons vivants, entre autres (Froese 1988 ; voir aussi Encadré 28).
La table OXYGEN contient des données sur la consommation d'oxygène des poissons, extraites d'expériences rapportées dans la littérature avec les facteurs connus ou qui affectent vraisemblablement le taux métabolique, particulièrement le poids corporel ; la température ; la salinité ; la concentration en oxygène ; le niveau d'activité ; la vitesse de nage ; et le facteur majeur de stress appliqué au sujet de l'étude. Comment [Commentaire] contient des détails expérimentaux supplémentaires, comme le nombre de spécimens, et d'autres informations. Les champs suivants fournissent des détails sur les facteurs listés ci-dessus.
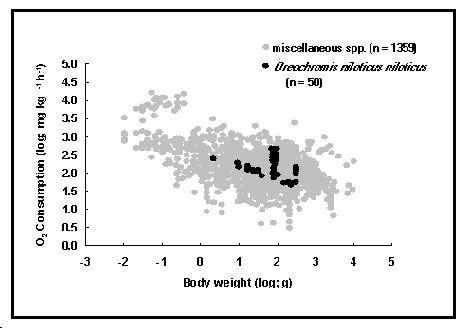
Fig. 45. Consommation relative d'oxygène chez Oreochromis niloticus niloticus comparée à diverses espèces. Noter la pente relativement bien descendante du nuage de points, et les séries verticales de valeurs obtenues par les différents stress appliqués.
Oxygen consumption [Consommation d'oxygène] : Indique la quantité d'oxygène utilisée par un poisson en mg·kg-1h-1. Si nécessaire, les données de la littérature ont été converties dans cette unité. De plus, un champ calculé a été inclus dans lequel les consommations d'oxygène mesurées à des températures comprises entre 5 et 30°C ont été calculées pour une température de 20°C en utilisant les multiplicateurs du tableau 3.3 de Winberg (1971).
Sex [Sexe] : Parmi les choix suivants : alevins ; juvénile ; femelle ; mâle ; mixte (pour mâle et femelle) ; non sexé (pour sexe inconnu).
Weight [Poids] : Indique le poids du spécimen testé en g. Le poids moyen est indiqué quand l'expérience a porté sur plus d'un individu à la fois.
Number [Nombre] : Indique le nombre total d'individus de l'expérience.
Temperature [Température] : Indique la température moyenne de l'eau, en °C, pendant l'expérience.
Salinity [Salinité] : La salinité moyenne en ‹ ppt › [‰], pendant l'expérience. Si la salinité n'avait pas été précisée, les valeurs de 35 ‰ et 0 ‰ ont été prises par défaut respectivement pour les espèces marines et d'eau douce. Pour les poissons diadromes, une valeur peut-être précisée dans le champ Comment [Commentaire]. Les valeurs erronées n'affecteront que légèrement la saturation en oxygène calculée.
Oxygen (mmHg) [Oxygène (mmHg)] : Indique la pression partielle moyenne d'oxygène en mmHg dans l'eau expérimentale. Quand cette valeur n'est pas précisée dans l'article original, elle est estimée à partir de la description de la méthode expérimentale, de la même manière que Thurston et Gehrke (1993). Ils proposent des corrections similaires pour la température et la salinité de l'eau expérimentale.
Oxygen (mg/l) [Oxygène (mg/l)] : Indique la concentration d'oxygène de l'eau expérimentale en mg · l-1. Si une valeur existe dans le champ précédent (Oxygène mmHg), la valeur en mg · l-1 est le résultat de la conversion suivante de la valeur en mmHg :
![]()
où ![]() est le
coefficient de Bunsen pour l'oxygène à la température et la
salinité données (Colt 1984).
est le
coefficient de Bunsen pour l'oxygène à la température et la
salinité données (Colt 1984).
100% oxygen saturation [Saturation d'oxygène 100%] : Indique, à titre de référence, la teneur maximale en oxygène (en mg · l-1) de l'eau à la température et à la salinité données.
Saturation% [% saturation] : Indique la teneur en oxygène réelle de l'eau expérimentale en pourcentage de la teneur maximale possible en oxygène. Les niveaux de saturation typiques sont autour de 90 %. Des valeurs inférieures à 70 % sont considérées comme ‹ hypoxie ›, les valeurs supérieures à 105 % comme ‹ supersaturation › (voir ‹ Stress Appliqué › ci-dessous).
Activity level [Niveau d'activité] : Permet de tenir compte de l'effet d'une activité sur le taux métabolique parmi les choix suivants : métabolisme standard (poisson au repos) ; métabolisme de routine (poisson spontanément actif) ; métabolisme actif (poisson nageant).
Swimming speed [Vitesse de nage] : Indique la vitesse de nage comme autre indice d'activité. La vitesse a été rapportée directement ou convertie en longueur du corps par seconde (LC/s) avec LC qui correspond généralement à la longueur totale ou à la longueur à la fourche.
Applied stress [stress appliqué] : Indique quel est le stress appliqué avant ou pendant l'expérience parmi les choix suivants : none specified [aucun spécifié] ; temperature [température] (variations ou valeurs extrêmes) ; photoperiod [photopériode] (durée exceptionnelle ou moment du jour de l'exposition à la lumière) ; feeding [alimentation] (pendant ou juste avant l'expérience) ; starvation [privation de nourriture] (sans nourriture offerte depuis plus de 24 h) ; toxins [toxines] ; hypoxia [hypoxie] (oxygène insuffisant) ; hypercapnia [hypercapnie] (teneur excessive de CO2 dans le sang quand l'individu est forcé de nager rapidement) ; salinity [salinité] (variations) ; high pH [pH élevé] ; low pH [pH faible] ; sedative [sédatif] ; transport [transport] ; other [autre]. Si le choix est ‹ autre ›, le stress est précisé dans Comment [Commentaire].
Utilisation
Les données de la table OXYGEN peuvent être utilisées pour tester des hypothèses sur les relations entre les différents stress et activités auxquels les poissons sont soumis, pour estimer la consommation d'énergie (nourriture) pour la modélisation trophique, et pour relier la croissance, la morphologie et le taux métabolique, entre autres.
Sources
La table OXYGEN contient probablement la plus grande compilation de données sur la consommation d'oxygène des poissons, avec près de 7 000 enregistrements pour environ 300 espèces extraites de plus de 400 références, entre autres Winberg (1960), Congleton (1974), Gorelova (1977), Marais (1978), Subrahamanyam (1980) et Neumann et al. (1981). 6 400 enregistrements proviennent de la base de données OXYREF compilée par Thurston et Gehrke (1993). Le reste a été ajouté par l'équipe FishBase.
Statut
La littérature originale a été consultée, les valeurs et les autres informations pertinentes rapportées et vérifiées. Cependant, cela a été fait pour peu de données jusqu'à présent. L'équipe FishBase continuera d'ajouter de nouveaux enregistrements et de vérifier les informations déjà saisies.
Comment y arriver
Cliquer sur le bouton Biology dans la vue SPECIES, puis sur le bouton Morphology & Physiology dans la vue BIOLOGY, et sur le bouton Metabolism dans la fenêtre MORPHOLOGY & PHYSIOLOGY. Double-cliquer sur une des lignes dans la vue LIST OF OXYGEN CONSUMPTION STUDIES affiche les informations relatives à l'étude désignée.
Remerciements
Nous sommes reconnaissants à R.V. Thurston et P.C. Gehrke pour avoir gracieusement confié la base de données OXYREF au projet FishBase pour élargir sa diffusion.
Références
Colt, J. 1984. Computation of dissolved gas concentrations in water as functions of temperature, salinity and pressure. Am. Fish. Soc. Spec. Publ. 14, 154 p.
Congleton, J.L. 1974. The respiratory response to asphyxia of Typhlogobius californiensis (Teleostei : Gobiidae) and some related gobies. Biol. Bull. 146 : 186-205.
Froese, R. 1988. Relationship between body weight and loading densities in fish transport using the plastic bag method. Aquacult. Fish. Manage. 19 : 275-281.
Gorelova, T.A. 1977. Respiration rate of the fry of some tropical fishes. Polskie Archiwum Hydrogiologii 24(Suppl.) : 447-453.
Marais, J.F.K. 1978. Routine oxygen consumption of Mugil cephalus, Liza dumerii and L. richardsoni at different temperatures and salinities. Mar. Biol. 50 : 9-16.
Neumann, D.A., J.M. O'Connor et J.A. Sherk, Jr. 1981. Oxygen consumption of white perch (Morone americana), striped bass (M. saxatilis) and spot (Leiostomus xanthurus). Comp. Biochem. Physiol. 69A : 467-478.
Subrahamanyam, C.B. 1980. Oxygen consumption of estuarine fish in relation to external tension. Comp. Biochem. Physiol. 67A : 129-133.
Thurston, R.V. et P.C. Gehrke. 1993. Respiratory oxygen requirements of fishes : description of OXYREF, a data file based on test results reported in the published literature, p. 95-108. In R.C. Russo et R.V. Thurston (éds) Fish physiology, toxicology, and water quality management. Proceedings of an International Symposium, Sacramento, California, USA, 18-19 September 1990. EPA/600AR-93/157.
Winberg, G.G. 1956. Rate of metabolism and food requirements of fishes. Minsk, USSR. Transl. Ser. Fish. Res. Board Can. 194, 1960.
Winberg, G.G. 1971. Methods for the estimation of production of aquatic animals. Academic Press, London. 175 p.
Armi Torres et Rainer Froese
L'immersion dans l'eau est l'approximation la plus proche de l'apesanteur. De plus, les téléostéens ont appris à échapper à l'influence de la gravité avec une moindre dépense d'énergie, grâce à leur vessie gazeuse. Le revers de l'existence sous l'eau est que tout mouvement entraîne des déplacements, sur les côtés, d'eau particulièrement dense et lourde. La physique et la physiologie de la nage des poissons sont résumées par Blake (1983), Webb (1984) et plus récemment par Videler (1993).
Ce chapitre décrit la manière dont nagent les poissons, ce qu'ils font d'un nombre surprenant de façons ; de plus, sont présentées les données disponibles sur leur vitesse de nage.
La classification des modes de nage des poissons adoptée ici est celle de Lindsey (1978) qui a beaucoup revu la littérature ancienne sur ce sujet. Cette classification consiste en deux niveaux, le premier (chiffres romains) décrivant ce qui peut être appelé le ‹ type › de nage, le second (puces) décrivant le ‹ mode › de nage proprement dit, c'est-à-dire :
I. Mouvements de la nageoire caudale et/ou du corps :
II. Ondulation des nageoires impaires et/ou pectorales :
III. Oscillations des nageoires impaires et/ou pectorales :
La série des modes du type I traduit une transition graduelle d'ondulations du corps entier (y compris le tronc) pour la propulsion (mode anguilliforme) aux seules oscillations de la nageoire caudale (modes thunniforme et ostraciiforme).
La classification des modes des types I à III traduit une transition graduelle des mouvements responsables de la force propulsive majeure, des ondulations vers les oscillations ; par conséquent, cette classification peut être aussi représentée par un graphique, dont l'axe tronc-nageoire caudale serait celui des ordonnées, et l'axe ondulation-oscillation celui des abscisses (voir Fig. 1 in Lindsey 1978).
La tâche d'attribuer un mode de nage à une espèce donnée contiendra toujours un élément subjectif, même sans tenir compte du fait que les poissons peuvent avoir deux modes de nage. Nous considérons ici seulement le mode dominant, à l'exclusion du mode de nage de fuite. Par exemple, les Scaridae passent d'un mode labriforme en période normale à un mode subcarangiforme quand ils fuient un danger présumé (observation personnelle des auteurs).
Aussi, les données de cette table (c'est-à-dire le choix des types et modes ci-dessus) peuvent-elles être révisées de temps en temps selon la progression de notre expérience de ce sujet et de sa littérature. Cependant, les données extraites de Lindsey (1978) (soit les espèces dont les champs MainRef. et Ref. pointent vers cette publication) et leurs extensions évidentes (par exemple, de Anguilla anguilla à tout Anguillidae, et à tout Anguilliformes) ne seront pas affectées par ces changements.
Noter aussi que cette table se rapporte seulement aux poissons juvéniles et adultes pour l'instant. Les larves, pour des raisons évidentes, ont un répertoire limité de types et de modes de nage.
L'indice de forme de la nageoire caudale d'une espèce correspond étroitement à son niveau moyen d'activité (Pauly 1989), et il est égal à :
A = h2 / s
où h est la hauteur de la nageoire caudale et s sa superficie (Fig. 46). Le bord proximal de la surface de la nageoire caudale est défini comme une ligne verticale droite traversant la portion la plus étroite du pédoncule caudal, c'est-à-dire que la portion du pédoncule entourée par la nageoire est considérée comme faisant partie de la surface de la nageoire caudale.
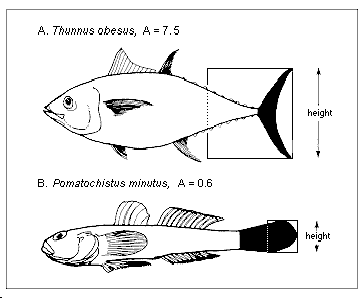
Fig. 46. Indice de forme (= h2/s où h = hauteur de la nageoire caudale ; s = superficie de la nageoire caudale) chez un poisson pélagique (A = 7,5) et chez un poisson benthique (B = 0,6). Noter la correspondance entre les indices de forme et les modes de vie.
Noter que cette définition de A diffère légèrement de celle présentée dans Pauly (1989), où seule la nageoire caudale a été utilisée pour calculer la valeur de s.
L'indice de forme peut être référencé, mais quand aucune référence n'est donnée, l'indice de forme a été calculé à partir de mesures faites par planimétrie sur une des illustrations dans FishBase ou d'une autre source facilement disponible.
Statut
La table SWIMMING contient les types et les modes de nage pour plus de 2 600 espèces.
La table SPEED contient 247 enregistrements de vitesse maximale de nage pour 79 espèces extraits de plus de 50 références, entre autres Bainbridge (1958, 1960) et Webb (1971), et de compilations comme Sambilay (1990). Un effort a été fait pour distinguer la nage ‹ soutenue › (nage maintenue plus de 3 minutes), de la nage ‹ explosive › (nage maintenue seulement quelques secondes) (voir Fig. 47), ainsi que d'autres modes de nage (Hammer 1995).

Fig. 47. Vitesse de nage en fonction de la longueur du corps chez les poissons. Noter que la vitesse de la nage ‹ explosive › est dix fois plus rapide que celle de la nage soutenue. Noter également qu'aucun poisson ne paraît nager à des vitesses inférieures à la limite de dépense d'énergie minimum par distance parcourue de Weihs (1973), dont la pente égale à 1 (impliquant une stricte proportionnalité entre vitesse et longueur) est pourtant la même que pour la courbe ci-dessus.
Les champs
Length [Longueur] : Indique la longueur du poisson en cm telle que citée dans la source. Le type de longueur utilisé est indiqué parmi les choix suivants : SL (LS, longueur standard) ; FL (LF, longueur à la fourche) ; TL (LT, longueur totale) ; BL (LC, longueur du corps, si la publication le mentionne ainsi, mais sans indication plus précise).
Swimming speed [Vitesse de nage] : Indique la vitesse du poisson par seconde telle que citée dans la source. Le type de longueur utilisé est indiqué comme pour le champ Length avec m/s en plus.
Speed [Vitesse] : Indique la vitesse convertie en m/s (voir le champ Derived values pour des valeurs dérivées).
Mode : Indique le mode de nage parmi les choix suivants : soutenue (nage à cette vitesse pendant une durée prolongée) ; explosive (nage à une vitesse maximale qui ne peut être maintenue que moins d'une minute) ; autre.
Comment [Commentaire] : Indique une longueur, un poids ou un mode de nage inclus dans les choix ci-dessus. Contient aussi toute conversion de type de longueur.
Derived values [Valeurs dérivées] : Indique des valeurs converties de différentes unités de vitesse en longueur standard par seconde (LS/s) et en mètre par seconde (m/s). La comparaison entre les performances de nage de poissons à nageoire caudale bifurquée, arrondie ou autre devient alors possible. Si elles sont disponibles, les valeurs des constantes a et b, utilisées dans l'équation suivante pour transformer la longueur à la fourche ou la longueur totale en longueur standard, sont indiquées :
LS = a + b ![]() L
L
où L est la longueur à la fourche ou la longueur totale citée. Si la transformation est fondée sur une mesure seule, en souhaitant que ce soit un adulte typique, le point d'intersection a est nul (voir aussi ‹ La table LENGTH-LENGTH ›, ce volume).
Comment y arriver
Cliquer sur le bouton Biology dans la vue SPECIES, puis sur le bouton Morphology & physiology dans la vue BIOLOGY et sur le bouton Swim. Type ou Swim. Speed dans la fenêtre MORPHOLOGY & PHYSIOLOGY.
Références
Bainbridge, R. 1958. The speed of swimming of fish as related to the size and to the frequency and amplitude of the tail beat. J. Exp. Biol. 35(1) : 109-133.
Bainbridge, R. 1960. Speed and stamina in three fish. J. Exp. Biol. 37(1) : 129-153.
Blake, R.W. 1983. Functional design and burst-and-coast swimming in fishes. Can. J. Zool. 61 : 2491-2494.
Hammer, C. 1995. Fatigue and exercise tests with fish. Comp. Biochem. Phsyiol. 112A(1) : 1-20.
Lindsey, C.C. 1978. Form, function, and locomotory habits in fish, p. 1-100. In W.S. Hoar et D.J. Randall (éds) Fish physiology. Vol. II. Academic Press, New York.
Pauly, D. 1989. A simple index of metabolic level in fishes. Fishbyte 7(1) : 22.
Sambilay, V.C., Jr. 1990. Interrelationships between swimming speed, caudal fin aspect ratio and body length of fishes. Fishbyte 8(3) : 16-20.
Videler, J.J. 1993. Fish swimming. Chapman and Hall, London. 260 p.
Webb, P.W. 1971. The swimming energetics of trout. Thrust and power output at cruising speed. J. Exp. Biol. 55 : 489-520.
Webb, P.W. 1984. Form and function in fish swimming. Sci. Am. 251 : 58-68.
Weihs, D. 1973. Optimal fish cruising speed. Nature 245 (Sept. 7) : 48-50.
Rainer Froese, Armi Torres, Crispina Binohlan et Daniel Pauly
Pour survivre, croître et se reproduire, les poissons ont besoin de nourriture et d'oxygène comme tout animal hétérotrophe. Cependant, bien qu'une littérature abondante existe sur la nourriture et les habitudes alimentaires des poissons (documentées dans plusieurs tables de FishBase), un faible nombre traite des organes et des processus qui permettent l'extraction d'énergie de cette nourriture.
Le processus essentiel est la respiration, et il est documenté, en partie, dans la table OXYGEN. Les organes importants, les branchies, sont traitées dans la table.
Cette table présente la plupart des mesures déjà publiées de superficie de branchies chez les poissons, car c'est cette superficie qui limite leur prise d'oxygène et donc leur taux métabolique et leur taux de croissance (Pauly 1979, 1981, 1994). La plupart de ces valeurs proviennent des compilations de Hughes et Morgan (1973), De Jager et Dekkers (1975), et Palzenberger et Pohla (1992).
Hughes (1984) discute quelques-uns des problèmes de mesure de la superficie des branchies et de leur interprétation ; ce travail devrait être consulté avant d'analyser les informations de la présente table. Pauly (1979, 1981, 1994) et Longhurst et Pauly (1987) présentent les éléments d'une théorie de la croissance des poissons dont certaines hypothèses peuvent être testées en utilisant des estimations de la superficie des branchies. Des utilisations pratiques de cette surface peuvent être envisagées dans des travaux écotoxicologiques et sur la pollution.
La figure 48 montre que la superficie des branchies augmente avec le poids corporel ; cependant, la pente inférieure à 1 de la courbe log-log implique que la superficie relative diminue avec la taille du corps.
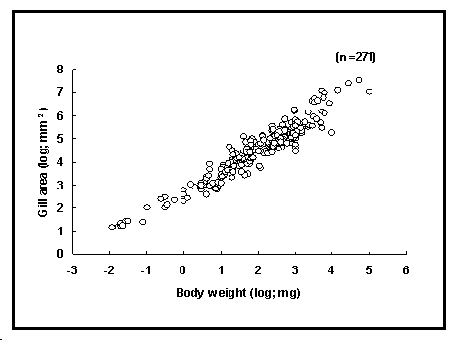
Fig. 48. Superficie des branchies en fonction du poids corporel (271 enregistrements pour 110 espèces).
La figure 49 de la courbe de la superficie relative des branchies en fonction du poids corporel en log-log le confirme, la pente étant égale à -0,2. Cependant, cette courbe masque des différences espèce-spécifiques qui se révèlent importantes dans les études des relations entre superficie des branchies et croissance (Pauly 1981).
Expliquer ces différences exige la prise en compte des modes de nage et/ou des indices de forme de la nageoire caudale. Nous espérons obtenir un graphique dans FishBase qui relie directement la performance de croissance et la superficie des branchies, et en prenant ces facteurs externes en considération.
De plus, le contenu de cette table sera mis à jour d'après les références appropriées, et vos suggestions sur ce sujet seront bienvenues.
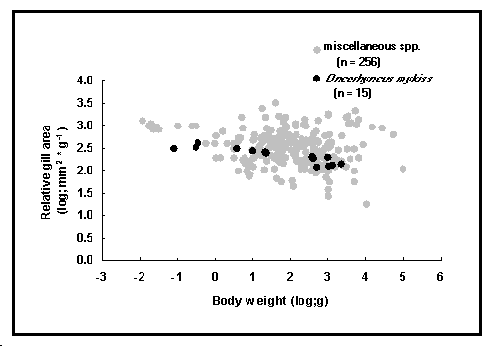
Fig. 49. Superficie des branchies de Oncorhynchus mykiss en fonction de son poids corporel (points noirs), comparée à divers poissons (points clairs).
Les champs
Gill area (in cm2) [Superficie des branchies (en cm2] : Champ-clé de cette table, chaque valeur doit toujours être reliée à une valeur de Body weight (in g) [Poids corporel (en g)].
Gill area/body weight (cm2/g) [Superficie des branchies/poids corporel (cm2/g)] : Champ calculé.
Blood/water distance [distance sang/eau] : Pour l'épaisseur de l'épithélium de la branchie (en nm).
Remarks [Remarques] : Contient tout commentaire méthodologique ou information pertinente.
Comment y arriver
Cliquer sur le bouton Biology dans la vue SPECIES, puis sur le bouton Morphology & physiology dans la vue BIOLOGY et sur le bouton Gill area dans la fenêtre MORPHOLOGY & PHYSIOLOGY. Double-cliquer sur une des lignes dans la vue LIST OF GILL AREA STUDIES affiche les informations relatives à l'étude désignée.
Remerciements
Je remercie le Professeur G.M. Hughes pour avoir répondu aimablement pendant toutes ces années à mes nombreuses questions au sujet des branchies des poissons.
Références
De Jager, S. et W.J. Dekkers. 1975. Relation between gill structure and activity in fish. Neth. J. Zool. 25 : 276-308.
Hughes, G.M. 1984. Measurement of gill area in fishes : practices and problems. J. Mar. Biol. Assoc. U.K. 64 : 637-655.
Hughes, G.M. et M. Morgan. 1973. The structure of fish gills in relation to their respiratory function. Biol. Rev. 48 : 419-475.
Longhurst, A. et D. Pauly. 1987. Ecology of tropical oceans. Academic Press, San Diego. 407 p.
Palzenberger, M. et H. Pohla. 1992. Gill surface area of water-breathing freshwater fish. Rev. Fish Biol. Fish. 2 : 187-216.
Pauly, D. 1979. Gill size and temperature as governing factors in fish growth : a generalization of von Bertalanffy's growth formula. Ber. Inst. Meereskd. Christian-Albrechts Univ. Kiel No. 63, xv + 156 p.
Pauly, D. 1981. The relationships between gill surface area and growth performance in fish : a generalization of von Bertalanffy's theory of growth. Meeresforsh./Rep. Mar. Res. 28(4) : 251-282.
Pauly, D. 1994. On the sex of fish and the gender of scientists. Chapman and Hall, London. 250 p.
Daniel Pauly
Cette table représente notre tentative pour incorporer dans FishBase les produits consommables à base de poissons.
Les champs
Dans son état actuel, la table comprend quatre groupes de champs :
Sources
La table PROCESSING contient 682 enregistrements pour 505 espèces de poissons, principalement extraits de Bykov (1983) dont l'ouvrage a servi de modèle à la conception de la table.
Rétrospectivement cependant, nous éprouvons le besoin de la repenser entièrement pour permettre :
Nous apprécierions des propositions de collègues intéressés à collaborer avec nous sur ce sujet, un développement qui rendrait FishBase utile à toute une nouvelle catégorie d'utilisateurs.
Comment y arriver
Cliquer sur le bouton Biology dans la vue SPECIES, puis sur le bouton Fish as food dans la vue BIOLOGY et sur le bouton Processing dans la fenêtre FISH AS FOOD. Double-cliquer sur une des lignes dans la vue LIST OF PROXIMATE ANALYSES affiche les informations relatives à la composition du produit désigné. Cliquer sur les boutons Weight proportions et Chemical composition pour afficher respectivement les pourcentages en poids et la composition chimique des différentes parties du corps.
Références
Bykov, V.P. 1983. Marine fishes : chemical composition and processing properties. Amerind Publishing Co., New Delhi. 333 p.
OECD. 1978. Multilingual dictionary of fish and fish products. Organization for Economic Co-operation and Development, Paris. 430 p.
Vinogradov, A.P. 1953. The elementary chemical composition of marine organisms. Memoirs, Sears Foundation for Marine Research II, New Haven. 647 p.
Daniel Pauly, Emily Capuli et Rainer Froese